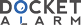

Pharmacological Profile of (2R-trans)-4-[1-[3,5-
`bis(Trifluoromethyl)benzoyl]-2-(phenylmethyl)-4-piperidinyl]-N-
`(2,6-dimethylphenyl)-1-acetamide (S)-Hydroxybutanedioate
`(R116301), an Orally and Centrally Active Neurokinin-1
`Receptor Antagonist
`A. A. H. P. MEGENS, D. ASHTON, J. C. A. VERMEIRE, P. C. M. VERMOTE, K. A. HENS, L. C. HILLEN,
`J. F. FRANSEN, M. MAHIEU, L. HEYLEN, J. E. LEYSEN, M. R. JURZAK, and F. JANSSENS
`Department of Discovery Research Johnson & Johnson Pharmaceutical Research and Development, Beerse, Belgium
`Received February 6, 2002; accepted April 9, 2002
`ABSTRACT
`In comparison with a series of reference compounds, (2R-trans)-
`4-[1-[3,5-bis(trifluoromethyl)benzoyl]-2-(phenylmethyl)-4-piperidi-
`nyl]-N-(2,6-dimethylphenyl)-1-acetamide (S)-Hydroxybutanedio-
`ate (R116301) was characterized as a specific, orally, and centrally
`active neurokinin-1 (NK
`1) receptor antagonist with subnanomolar
`affinity for the human NK1 receptor (Ki: 0.45 nM) and over 200-fold
`selectivity toward NK2 and NK3 receptors. R116301 inhibited sub-
`stance P (SP)-induced peripheral effects (skin reactions and
`plasma extravasation in guinea pigs) and a central effect (thump-
`ing in gerbils) at low doses (0.08–0.16 mg/kg, s.c. or i.p.), reflect-
`ing its high potency as an NK
`1 receptor antagonist and excellent
`brain disposition. Higher doses blocked various emetic stimuli in
`ferrets, cats, and dogs (ED
`50 values: 3.2 mg/kg, s.c.; 0.72–2.5
`mg/kg, p.o.). Even higher doses (11–25 mg/kg, s.c.) were required
`in mice (capsaicin-induced ear edema) and rats (SP-induced ex-
`travasation and salivation), consistent with lower affinity for the
`rodent NK
`1 receptor and known species differences in NK1 re-
`ceptor interactions. R116301 inhibited the ocular discharge (0.034
`mg/kg) but not the dyspnoea, lethality, or cough (/H1102240 mg/kg, s.c.)
`induced by [
`/H9252ALA8]-neurokinin A (NKA) (4–10) in guinea pigs,
`attesting to NK 1 over NK2 selectivity. R116301 did not affect
`senktide-induced miosis (/H110225 mg/kg, s.c.) in rabbits, confirming
`the absence of an interaction with the NK3 receptor. R116301 was
`inactive in guinea pigs against skin reactions induced by hista-
`mine, platelet-aggregating factor, bradykinin, or Ascaris allergens
`(/H1102210 mg/kg, s.c.). In all species, R116301 showed excellent oral
`over parenteral activity (ratio, 0.22–2.7) and a relatively long dura-
`tion (6.5–16 h, p.o.). The data attest to the specificity and sensi-
`tivity of the animal models and support a role of NK
`1 receptors in
`various diseases.
`Tachykinins belong to a family of short peptides that are
`widely distributed in the mammalian central and peripheral
`nervous system (Lundberg, 1995; Maggi, 1995; Bertrand and
`Geppetti, 1996). They share the common C-terminal se-
`quence Phe-Xaa-Gly-Leu-Met-NH
`2. Tachykinins released
`from peripheral sensory nerve endings are believed to be
`involved in neurogenic inflammation. In the spinal cord/cen-
`tral nervous system, tachykinins may play a role in pain
`transmission/perception and in some autonomic reflexes and
`behaviors. The three major tachykinins are substance P (SP),
`neurokinin (NK) A and NKB with preferential affinity for
`three distinct receptor subtypes, termed NK
`1,N K2, and NK3,
`respectively. However, functional studies on cloned receptors
`suggest strong functional cross-interaction between the three
`Article, publication date, and citation information can be found at
`http://jpet.aspetjournals.org.
`DOI: 10.1124/jpet.102.034348.
`ABBREVIATIONS:SP, substance P; NK, neurokinin; NKA, neurokinin A; R116301, (2R-trans)-4-[1-[3,5-bis(trifluoromethyl)benzoyl]-2-(phenylmethyl)-4-
`piperidinyl]-N-(2,6-dimethylphenyl)-1-acetamide ( S)-hydroxybutanedioate; CGP49823, (2 R,4S)-2-benzyl-1-(3,5-dimethylbenzoyl)-4-(quinolin-4-
`ylmethylamino)piperidine; CP-96345, (2S,3S)-cis-2-(diphenylmethyl)-N-((2-methoxyphenyl)-methyl)-1-azabicyclo(2.2.2.)-octan-3-amine; CP-99994, (2S,3S)-3-
`(2-methoxybenzylamino)-2-phenylpiperidine; GR-203040, (2S,3S)-(2-methoxy-5-tetrazol-1-ylbenzyl)(2-phenylpiperidin-3-yl)amine; MK-869 or L-754030,
`aprepitant; L-760,735, 2-(R)-(1-(R)-3,5-bis(trifluoromethyl)phenyl)ethoxy)-4-(5-(dimethylaminomethyl)-1,2,3-trioazol-4-yl)methyl-3-(5)-phenyl)morpholine; RP-
`67580, (3aR,7aR)-7,7-diphenyl-2-[1-imino-2-(2-methoxyphenyl)ethyl]perhydroisoindol-4-one; SDZ-NKT-343, 2-nitrophenylcarbamoyl-(S)-prolyl-(S)-3-(2-
`naphthyl)alanyl-N-benzyl-N-methylamide; SR-140333, nolpitantium; SR-48968, saredutant; SR-142801, osanetant; Y-24180, (/H11006)-4-(2-chlorophenyl)-2-[2-(4-
`isobutylphenyl)ethyl]-6,9-dimethyl-6H-thieno[3,2-f][1,2,4]triazolo[4,3-a][1,4]diazepine; LY255582, (3R,4R)-3,4-dimethyl-1-[(3S)-3-hydroxy-3-cyclohexyl-propyl]-
`4-(3-hydroxyphenyl)piperidine; PAF, platelet-aggregating factor; 5-HT, serotonin; MDL-103392, 4-piperidinecarboxamide, 1-[2-[3-(3,4-dichlorophenyl)-1-(3,4,5-
`trimethoxybenzoyl)-3-pryrrolidinyl]ethyl]-4-phenyl; MDL-105212, (3R)-MDL-103392.
`0022-3565/02/3022-696–709$7.00
`THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 302, No. 2
`Copyright © 2002 by The American Society for Pharmacology and Experimental Therapeutics 34348/996964
`JPET 302:696–709, 2002 Printed in U.S.A.
`696
`HELSINN EXHIBIT 2066
`Azurity Pharmaceuticals, Inc. v. Helsinn Healthcare S.A.
`IPR2025-00948
`Page 1 of 14
`
`
`
`
`
`
`
`tachykinins and their corresponding receptors (Maggi and
`Schwartz, 1997). The tachykinins are involved in emesis,
`anxiety states (stress-related), smooth muscle contraction,
`inflammation, and nociception/pain perception, although
`clinically relevant involvement of SP, particularly for the last
`items, is questionable (Hill, 2000).
`R116301 is a new NK
`1 receptor antagonist (Fig. 1 ). The
`present study reports on its pharmacological profile in sev-
`eral NK receptor-related models in various species. Species
`differences in the structure of NK
`1 receptors are responsible
`for species-related potency differences of NK1 receptor antag-
`onists (Maggi, 1995). The human NK1 receptor closely resem-
`bles the NK 1 receptor of guinea pigs and gerbils but differs
`markedly from the NK1 receptor of rodents. Therefore, most
`tests are performed in guinea pigs and gerbils. Attention is
`paid to potency, selectivity, specificity, onset and duration of
`action, and species differences. R116301 is compared with
`the following available NK
`1 receptor antagonists (Fig. 1 for
`chemical structures): CGP49823 (Vassout et al., 1994), CP-
`96345 (Snider et al., 1991), CP-99994 (Piedimonte et al.,
`1993), GR-203040 (Ward et al., 1995), MDL-103392 (race-
`mate of the active enantiomer MDL-105212; Kudlacz et al.,
`1996), aprepitant (MK-869 or L-754030; Kramer et al., 1998;
`Rupniak and Kramer, 1999), L-760735 (McAllister et al.,
`Fig. 1. Chemical structure of R116301 [(S)-hydroxybutanedioate salt] and the reference NK 1 receptor antagonists.
`Pharmacology of R116301 697
`Page 2 of 14
`
`
`
`
`
`
`
`1999), RP-67580 (Garret et al., 1991), SDZ-NKT-343 (Wal-
`pole et al., 1998), nolpitantium (SR-140333; Emonds-Alt et
`al., 1993). The testing of some compounds was restricted by
`limited availability. The sensitivity, reliability, and specific-
`ity of the various test models is evaluated based on the
`results obtained with the test compounds. A preliminary
`article on R116301 has been presented in poster format
`(Jurzak et al., 2000). A patent application has been filed
`(Janssens et al., 1997).
`Materials and Methods
`Animals
`Swiss mice (Janssen Pharmaceutica, Beerse, Belgium), Wistar
`rats (Janssen Pharmaceutica), Mongolian CRW gerbils ( Meriones
`unguiculatus; Charles River Breeding Laboratories, Inc., Sulzteld,
`Germany), Dunkin-Hartley-Purbright guinea pigs (Janssen Phar-
`maceutica or Charles River Breeding Laboratories, Inc.), Fish ferrets
`(Harlan CPB, Horst, The Netherlands), New Zealand white rabbits
`(Broekman Institute, Someren, The Netherlands), cats (Broekman
`Institute), and Beagle dogs (Janssen Pharmaceutica or Harlan CPB)
`were used. They were fasted overnight (tap water remained avail-
`able ad libitum) and housed under standard laboratory conditions
`(21 /H110062°C; 65 /H1100615% relative humidity; light/dark cycle set at 12 h).
`During the test period, they were housed in individual cages. The
`ferrets, cats, and dogs were used more than once, with an intertrial
`interval of at least 1 week. All animal studies were approved by the
`local Ethical Committee in compliance with the Declaration of Hel-
`sinki.
`Chemicals and Test Compounds
`R116301 was dissolved up to 2.5 mg/ml in 10% hydroxypropyl- /H9252-
`cyclodextrin. Higher doses were prepared as suspensions in 1% poly-
`sorbate 80 in distilled water. The preparations were stored at room
`temperature in closed containers protected from light. They were
`studied at various time intervals after subcutaneous (10 ml/kg for
`mice, rats, and guinea pigs; 1.0 ml/kg for rabbits; 0.5 ml/kg for cats
`and dogs), oral (10 ml/kg for mice, rats, and guinea pigs; 1.0 ml/kg for
`rabbits; 0.5 ml/kg for cats and dogs), or intravenous administration
`(10 ml/kg for mice; 2 ml/kg for rats; 0.5 ml/kg for rabbits, cats, and
`dogs). All doses were expressed in milligram base equivalents per
`kilogram body weight. With the exception of aprepitant (L-754030 or
`MK-869; synthesized by and obtained from Johnson & Johnson
`Pharmaceutical Research Institute and Development, Springhouse,
`NJ) and L-760735 (synthesized by our Department of Medicinal
`Chemistry), the reference compounds were kindly provided by the
`companies of origin: CGP49823 and SDZ-NKT-343 (Novartis, Basel,
`Switzerland); CP-96345 and CP-99994 (Pfizer, Sandwich, Kent, UK);
`GR-203040
`(GlaxoSmithKline, Uxbridge, Middlesex, UK); MDL-103392 (Aven-
`tis, Strasbourg, France); RP-67580 (Aventis); saredutant/SR-48968,
`nolpitantium/SR-140333, and osanetant/SR-142801 (SANOFI Re-
`search Center, Montpellier, France). For all these compounds with
`exception of SR-140333 and SDZ-NKT-343, some batches were syn-
`thesized in our Department of Medicinal Chemistry. The origin of
`the other chemicals is indicated in parentheses after each name:
`substance P (Sigma-Aldrich, St. Louis, MO), [
`/H9252ALA8]-neurokinin A
`(4–10) (NovaBiochem, Laufelfingen, Switzerland), capsaicin (Sigma-
`Aldrich), PAF (Sigma-Aldrich), bradykinin (Sigma-Aldrich), hista-
`mine (Sigma-Aldrich), and Evans blue dye (Sigma-Aldrich; Direct
`blue 53; dye content approximately 85%). Ascaris suum worms were
`obtained from freshly slaughtered pigs. A batch of about 20 ml of
`perienteric fluid was centrifuged; the clear supernatant was divided
`in 0.1-ml portions and stored at /H1100218°C. The injected solutions of
`Ascaris coeloma fluid were fresh 1:16 dilutions in 0.9% NaCl.
`Pharmacological Tests
`In Vitro Receptor Binding. Membrane preparation from cells.
`Chinese hamster ovary cells expressing human (h)NK1,N K2 or NK3
`receptors were grown in Dulbecco’s modified Eagle’s medium/HAM’s
`F-12 medium containing 10% fetal calf serum and antibiotics. Twen-
`ty-four hours after stimulation with 5 mM Na-butyrate to enhance
`expression levels, the cells were collected from plates using a rubber
`scraper and suspended in 50 mM Tris-HCl, pH 7.4. They were
`centrifuged at 23,500g for 10 min in a Sorvall-RC 5B centrifuge
`(DuPont Instruments, Meyvis, Belgium). The pellets were homoge-
`nized in 5 mM Tris-HCl, pH 7.4, using an Ultra-Turrax homogenizer
`(Janke & Kunkel IKA Labortechnik, Staufen im Breisgau, Germany)
`and centrifuged at 30,000g for 20 min. The final pellet was sus-
`pended in 50 mM Tris-HCl, pH 7.4. The membranes were frozen in
`1-ml aliquots /H1100270°C. Before use, vials were thawed and rehomog-
`enized in incubation buffer B (50 mM Tris-HCl, pH 7.4, containing 2
`mM MgCl
`2, 1 mM EGTA, and 0.1% bovine serum albumin). The
`protein concentration was determined after using a Bradford kit
`from Bio-Rad (Hercules, CA).
`Membrane preparation from brain tissue. To prepare membranes
`for [
`3H]substance P binding from guinea pigs, gerbils, ferrets, and
`rats, animals were killed by decapitation, and the forebrains were
`dissected. The tissue was homogenized in 50 mM Tris-HCl, pH 7.4,
`using an Ultra-Turrax homogenizer. The homogenates were centri-
`fuged at 23,500g for 10 min at 0 –4°C in a Sorvall-RC 5B centrifuge.
`The pellets were washed twice in 50 mM Tris-HCl, pH 7.4, by
`resuspension with a dual-homogenizer (Kimble Kontes, Vineland,
`NJ) and centrifugation. After the last wash, the pellets were sus-
`pended in incubation buffer A [50 mM Tris-HCl, pH 7.4, containing
`2 mM MgCl
`2, 1 mM EGTA, 1 mM o-phenantrolin, and 0.1% bovine
`serum albumin at a dilution of 25 volumes/original wet weight of
`tissue (v/w)]. The final pellet of rat forebrain was suspended in
`incubation buffer B at a dilution of 40 v/w. Guinea pig, gerbil, and rat
`forebrain membranes were always freshly prepared. For ferret fore-
`brain, the membranes were prepared in 50 mM Tris-HCl, pH 7.4,
`and stored at a dilution of 5 v/w at /H1100270°C. Before use, these mem-
`branes were thawed and further diluted until dilution of 25 v/w in
`incubation buffer A.
`[
`3H]Substance P binding to NK1 receptors and data analysis. The
`membrane homogenates were incubated for 20 min at 25°C with 0.5
`nM [
`3H]SP (for rat membranes 1 nM was used) in a total volume of
`0.5 ml. Specific binding of the radioligand was distinguished from
`nonspecific binding by addition of unlabeled SP to a final concentra-
`tion of 10
`/H110027M. The incubation was stopped by the addition of 5 ml of
`ice-cold 50 mM Tris-HCl buffer, pH 7.4, followed by rapid filtration
`over Whatman GF/B glass fiber filters (Maidstone, Kent, UK) (pre-
`soaked in 0.1% polyethylenimine for 1 h) using a 40-well filtration
`unit. The filters were washed twice with ice-cold buffer to remove
`nonbound radioactivity and placed in plastic miniature vials. After a
`24-h incubation with 2 ml of Ultima Gold scintillation cocktail, vials
`were vigorously shaken, and the radioactivity was counted in a
`Packard Tri-Carb 1500 CA liquid scintillation analyser (Packard
`BioScience, Meriden, CT).
`R116301 was also investigated in various other in vitro binding
`assays using membrane preparations of animal tissue or membranes
`of cell lines transfected with cloned human receptors. Brain, periph-
`eral organs, blood of animal or human origin, or permanent cell lines
`were used as tissue sources. The procedures for membrane prepara-
`tions and references to the various receptor binding models were
`described previously (Briejer et al., 2001).
`Substance P-Induced Plasma Protein Extravasation in
`Guinea Pigs. SP (2
`/H9262g/kg, i.v.) and Evans blue dye (30 mg/kg, i.v.)
`were injected simultaneously (one solution; 4 ml/kg) into the femoral
`artery of guinea pigs of both sexes (325– 425 g) at predefined, loga-
`rithmically spaced, time intervals (1, 2, 4, 8, 16 or 32 h) after
`pretreatment with test compound or solvent. Up to 10 min after
`challenge, the animals were scored by visual inspection for blue
`698 Megens et al.
`Page 3 of 14
`
`
`
`
`
`
`
`coloring (0, 1, 2, or 3) of the nose, the forepaws, and the conjunctiva.
`The criterion for drug-induced inhibition of plasma extravasation:
`score /H110212 (occurrence in 0.2, 0.0, and 0.0% for blue coloring of the
`nose, paws, and conjunctiva, respectively, in saline-treated controls;
`n /H11005500).
`Capsaicin-Induced Ear Edema in Mice. Ear edema was in-
`duced by local application of capsaicin (250
`/H9262g) on the ear of female
`mice (20 –26 g) pretreated with test compound or solvent (modified
`after Inoue et al., 1996). The capsaicin was dissolved in acetone at a
`concentration of 12.5 mg/ml and applied in a volume of 10
`/H9262l both on
`the ventral and dorsal side of the left ear. Pure acetone was applied
`in a volume of 10
`/H9262l both on the ventral and dorsal side of the right
`ear, which served as a control measure. Immediately after the cap-
`saicin challenge, Evans blue dye (7.5 mg/ml in 0.9% NaCl) was
`injected intravenously in a volume of 10 ml/kg, corresponding to a
`dose of 75 mg/kg. Thirty minutes after the capsaicin application, ear
`edema was scored (0, 1, 2, or 3) as the blue coloring of the left ear.
`Criterion for drug-induced inhibition: score /H110212 (never observed in
`controls; n /H1100562).
`Substance P-Induced Salivation and Plasma Protein Ex-
`travasation in Rats. Plasma protein extravasation and salivation
`were induced by injection of SP (2
`/H9262g/4 ml/kg, i.v.) in the tail vein of
`female rats (200 –240 g) that were anesthetized with pentobarbital
`(40 mg/kg, i.p.; 10 min before the SP challenge) and placed on a
`heating pad to maintain body temperature [modified after Snider et
`al. (1991); Robineau et al. (1995)]. Evans blue dye (30 mg/kg, i.v.) was
`injected simultaneously with SP (one solution; 4 ml/kg). Test com-
`pound or solvent was administered at a predefined time interval
`before SP injection. From 0 through 4 min after the SP challenge,
`salivation was measured by placing cotton swabs in the rat ’s mouth
`at 2-min intervals and quantifying the amount of saliva secreted by
`the difference in the weight of the two cotton swabs before and after
`the collection period. Five minutes after challenge, the animals were
`scored for blue coloring (0, 1, 2, or 3) of the nose and the paws. Thirty
`minutes after challenge, the animals were sacrificed by CO
`2 asphyx-
`iation and scored (0, 1, 2, or 3) for blue coloring of the trachea and
`urinary bladder. Criteria for drug-induced inhibition: /H11021100 mg of
`saliva for inhibition of salivation (2.0% in controls;n /H11022100); score /H110212
`for inhibition of plasma extravasation (0, 0, 1.0, and 1.0 for blue
`coloring of the nose, paws, trachea, and urinary bladder, respec-
`tively, in controls).
`Skin Reaction Test in Guinea Pigs: SP, Histamine, PAF,
`Bradykinin, and Ascaris Allergens. Five dorsal skin sites of
`guinea pigs of both sexes (300 –500 g) were injected intradermally
`(0.05 ml) with saline containing SP (0.05
`/H9262g), histamine (0.25 /H9262g),
`PAF (0.00625 /H9262g), bradykinin (0.1 /H9262g), and Ascaris allergens (1/16
`diluted with saline). Immediately thereafter, the animals were chal-
`lenged i.v. with Evans blue dye (30 mg/kg; 7.5 mg/ml, 4 ml/kg) and
`sacrificed 30 min later. The intensity of the blue-colored skin reac-
`tions was scored by two independent observers in comparison with
`standard reactions. The scoring system was maximal (4), pronounced
`(3), moderate (2), slight (1), and no difference (0) with surrounding
`skin. The scores of the two observers were summed for further
`evaluation. Test compound or solvent was administered at a pre-
`defined interval before induction of the skin reactions. Criteria for
`drug-induced effects were: 1) SP reactions: score /H110217 for inhibition,
`score /H110215 for pronounced inhibition, and score/H110213 for blockade (oc-
`curring in 3.6, 0.0, and 0.0%, respectively, of solvent-treated control
`guinea pigs; n /H1100557); 2) histamine reactions: score /H110217 for inhibition
`and score /H110213 for blockade (occurring in 3.6% and 0.0%, respectively,
`of the controls); 3) PAF reactions: score /H110217 for inhibition and score
`/H110213 for blockade (both occurring in 0.0% of the controls); and 4)
`bradykinin reactions: score /H110217 for inhibition and score/H110213 for block-
`ade (both occurring in 0.0% of the controls); and 5) Ascaris allergens
`reactions: score /H110213 for blockade (occurring in 5.4% of the controls).
`SP-Induced Thumping in Gerbils. Male gerbils (50–70 g) were
`prepared for intracerebral injection under anesthesia (exposure to
`4% isoflurane in 70% N
`2O /H1100130% O2 during 1 min) by making an
`incision in the skin above the cranium (modified after Bristow and
`Young, 1994 and Rupniak et al., 1997). SP (400 ng in 2 /H9262l) was
`rapidly injected into the cerebral ventricles after advancing a Ham-
`ilton needle 3.5 mm below a point 2 mm anterior to bregma and 1 mm
`lateral to the midline; the needle was removed 10 s later. The
`animals were observed for thumping behavior during the following 5
`min. Thumping was defined as a sharp downward movement of the
`hind-paws producing a characteristic rhythmic tapping sound. The
`onset of the SP-induced thumping, the total number of thumps, and
`the total thumping time (seconds) were registered. The animals were
`then immediately euthanized with CO
`2. For the present purpose,
`absence of thumps during the 5-min observation period was adopted
`as an all-or-nothing criterion for inhibition of the SP-induced thump-
`ing (1.2% false positive controls; n /H11005625).
`[
`/H9252ALA8]-NKA (4–10)-Induced Lethality in Guinea Pigs. The
`following phenomena were scored or noted after injection of [/H9252ALA8]-
`neurokinin A (4 –10) (50 /H9262g/4 ml/kg; i.v.) into the vena saphena of
`female guinea pigs (300 –500 g) pretreated with test compound or
`solvent: cough (intensity scores: 0, 1, or 2), white ocular discharge
`(scores: 0, 1, or 2), dyspnoea (score 1 /H11005cyanosis, score 2 /H11005dyspnoea,
`score 3 /H11005loss of righting reflex and/or clonic convulsions), and
`lethality (the survival time up to 60 min after [
`/H9252ALA8]-NKA (4 –10)
`challenge is noted) (modified after Robineau et al., 1995). [ /H9252ALA8]-
`neurokinin A (4 –10) is a metabolism-resistant analog of neuroki-
`nin-A with a high degree of selectivity for the NK 2 receptor (Rovero
`et al., 1989). The various phenomena generally occurred within 3 to
`5 min after the [
`/H9252ALA8]-NKA (4–10) challenge. The following all-or-
`nothing criteria were adopted for the determination of the ED 50
`values of the test compounds: score /H110050 for antagonism of cough
`(occurrence: 0.0% false positives in the control population; n /H1100596),
`score /H110050 for antagonism of ocular discharge (2.1% false positives),
`score /H110212 for protection from dyspnoea (1.0% false positives), and
`survival time /H1102260 min for protection from lethality (5.2% false pos-
`itives).
`Emesis in Ferrets, Cats, and Dogs. Vomiting or retching was
`induced by the peripherally selective opioid loperamide (0.31 mg/kg,
`s.c.), by ipecac syrup (1 ml/kg, p.o.; from a commercial source and
`containing ether-soluble alkaloids of ipecac in a concentration of 1.4
`mg/ml), by the dopamine agonist apomorphine (0.31 mg/kg, s.c.), or
`by the
`/H92512-adrenoceptor agonist xylazine (1.25 mg/kg, s.c.) in male
`ferrets (1–3 kg), cats (3–7 kg), or dogs (5–20 kg) pretreated with test
`compound or solvent. The number of retches or vomits was counted
`over a 1 h-period or (in the case of ipecac syrup-induced emesis in
`cats) a 3 h-period starting immediately after the emetic challenge.
`Loperamide-induced retching in ferrets. In control animals pre-
`treated with solvent, loperamide (0.31 mg/kg, s.c.) induced pro-
`nounced retching (mean /H11006S.D.: 95 /H1100639 counts; n /H11005529) and, to a
`lesser extent, vomiting (5 /H110064). All-or-nothing criteria for drug-
`induced effects on retching: /H1102120 retches for inhibition (2.0% false
`positives) and 0 retches for blockade (0% false positives).
`Ipecac syrup-induced retching in ferrets. Oral administration of
`ipecac syrup induced pronounced retching (mean /H11006S.D.: 76 /H1100636
`counts) and, to a lesser extent, vomiting (mean /H11006S.E.: 5.6 /H110062.3
`counts; n /H1100598) in control animals. Absence of retching or vomiting
`never occurred in these control animals and was used as criterion for
`antiemetic activity.
`Apomorphine-induced retching in ferrets.Injection of apomorphine
`resulted in pronounced retching in control ferrets (mean/H11006S.D.: 63 /H11006
`29 counts; n /H1100592) and, to a lesser extent, vomiting (mean /H11006S.D.:
`3.1 /H110062.0 counts). Absence of retching never occurred in these control
`animals and was used as criterion for antiemetic activity. Vomiting
`was absent in 13% of the control animals.
`Apomorphine-induced vomiting in dogs. Apomorphine induced
`consistent vomiting in all control dogs (mean /H11006S.D.: 10 /H110065 vomits;
`n /H11005195). Criteria for drug-induced protection: inhibition of emesis:
`/H110212 emetic bouts (occurring in 0.12% of controls); blockade of emesis:
`complete absence of emesis (not observed in controls).
`Xylazine-induced retching in cats. Control cats (n /H11005113) retched
`Pharmacology of R116301
`699
`Page 4 of 14
`
`
`
`
`
`
`
`(mean /H11006S.D.: 41 /H1100618 counts) and vomited (mean /H11006S.D.: 2.9 /H110061.8
`counts) after injection of xylazine. Absence of retching or vomiting
`never occurred and was used as criterion for antiemetic activity.
`Ipecac syrup-induced vomiting in cats. Oral administration of ip-
`ecac syrup induced vomiting and retching during a 3-h observation
`period in control cats pretreated with distilled water (n /H1100530). For
`practical reasons, only the number of emetic bouts was counted,
`which remained relatively low (mean /H11006S.E.: 4.5 /H110060.6 counts).
`Absence of vomiting never occurred and was used as the criterion for
`antiemetic activity.
`Senktide-Induced Miosis in Rabbits. Senktide (12.5
`/H9262g/0.5
`ml/kg, i.v.)-induced miosis was evaluated just before and 15 min
`after challenge in male New Zealand white rabbits (approximately 1
`kg) pretreated with test compound or solvent (1.0 ml/kg) (modified
`after Medhurst et al., 1997). The senktide was injected into an ear
`vein. The left pupil of each rabbit was measured under normal
`ambient lighting with a comparing reticule (scaled to 0.5 mm). In
`solvent-pretreated control rabbits (n /H1100573), pupil diameter declined
`from 6.0 /H110060.5 mm immediately before to 2.7 /H110060.7 mm 15 min after
`the senktide challenge. At this 15-min time interval after senktide
`injection, only two of the control rabbits (2.7%) showed a pupil
`diameter /H113505.0 mm, which was adopted as all-or-nothing criterion for
`significant inhibition of the senktide-induced miosis by test com-
`pounds.
`General Procedure and Statistics
`In Vitro Binding Studies. Sigmoidal inhibition curves were
`calculated by computerized nonlinear regression analyses according
`to Oestreicher and Pinto. pIC
`50 values (/H11002log concentration producing
`50% inhibition of specific [ 3H]SP binding) were derived. Inhibition
`equilibrium constants (K i values) were calculated according to Ki
`(nanomolar) /H11005IC50/(1 /H11001C/Kd), where C is the concentration and Kd
`the equilibrium dissociation constant of the [ 3H]SP.
`In Vivo Functional Studies.All experiments were performed by
`unbiased trained technicians using coded solutions. Doses were se-
`lected from the geometrical series 0.00063–0.00125–0.0025. . . 40.0–
`80.0–160 mg/kg in such a way that at least three doses covered the
`0 to 100% effect range of the dose-response curve. Each dose group
`consisted of five animals, which were tested in separate daily exper-
`imental sessions (including solvent-treated control animals) to ac-
`count for day-to-day variability and to minimize systematic errors.
`Control injections of solvent were included in each experimental
`session. Based on an analysis of a frequency distribution of a large
`series of the historical control data, all-or-nothing criteria for stim-
`ulation or inhibition were defined as a change of the measured or
`scored variable to values that never or almost never (in less than 5%)
`occurred in the control population. With the help of the thus defined
`all-or-nothing criteria, graded data were transformed to categorical
`data which generated per dose level the number of animals in which
`the intensity of a particular phenomenon was higher (stimulation) or
`lower (inhibition or blockade) than in the control animals. On the
`basis of the thus obtained dose-response relations, ED
`50 values and
`corresponding 95% confidence limits were calculated by probit anal-
`ysis according to the method of Finney (1962) for categorical data.
`Results
`In Vitro Receptor Binding Profile
`R116301 showed subnanomolar affinity for the human
`NK1 receptor (pIC50 /H11006S.D.: 8.53 /H110060.06; Ki: 0.45 nM; n /H110054
`replicates), with 230- and 1600-fold selectivity regarding
`binding to the human NK3 (pIC50 /H11006S.D.: 6.22 /H110060.14; Ki: 104
`nM; n /H110053) and NK2 (pIC50 /H11006S.D.: 5.87 /H110060.36; Ki: 711 nM;
`n /H110054) receptors, respectively. R116301 showed very low
`affinity for human serotonin 5-HT 2B (pIC50 /H11006S.D.: 5.68 /H11006
`0.15; Ki: 928 nM;n /H110052) and 5-ht5a (pIC50 /H11006S.D.: 5.16 /H110060.04;
`Ki: 3544 nM; n /H110052) receptors and for rat Ca2/H11001ligand binding
`sites (pIC50 /H11006S.D.: 5.45 /H110060.15; Ki: 2454 nM; n /H110053) and Na/H11001
`channels (pIC50 /H11006S.D.: 5.60 /H110060.17; Ki: 2526 nM; n /H110053). Up
`to 10 /H9262M, the compound had no affinity for a wide range of
`other binding sites, including adrenergic receptors (human
`/H92511A-, /H92512A-, /H92512B-, /H92512C-, /H92521-, /H92522-, /H92523-), dopaminergic (rat D 1;
`human D 2L,D 3, and D 4), serotonergic (human 5-HT 1A,
`5-HT1B, 5-HT 1D, 5-ht 1e, 5-ht 1f, 5-HT 2A, 5-HT 2C, 5-HT 3,
`5-HT4b, and 5-HT 7), human histamine H 1, rat cholinergic
`muscarinic, transporters (rat DA, NE; human 5-HT, GlyT 1,
`and GlyT 2), aspartate (rat NMDA-MK801, NMDA-glycine,
`and AMPA), opioid (human /H9262, /H9254; guinea pig /H9260), human halo-
`peridol-sensitive /H92681, peptide receptors (rat CCK A; human
`CCKB, bradykinin-B2, VIP). Since tachykinin receptors are
`known to exhibit strong species dependence in pharmacology,
`the NK1 affinity of R116301 was investigated on tissue mem-
`brane preparations from forebrains of guinea pigs, gerbils,
`ferrets, and rats. Relative to its affinity for the human NK
`1
`receptor, R116301 showed about 10 times lower affinity for
`the gerbil, ferret, and guinea pig NK
`1 receptors (Ki: 6.4, 8.3,
`and 13 nM, respectively) and 200 times lower affinity for the
`rat NK
`1 receptor (K i: 98 nM). These results show a high
`affinity, selectivity, and specificity of R116301 for the human
`NK
`1 receptor and a species preference as generally observed
`for other NK1 receptor antagonists as well.
`Fig. 2. Individual scores (ranging from 0 to 3) for SP-induced blue
`coloring of the nose, forepaws, and conjunctiva of guinea pigs 1 h after s.c.
`pretreatment with saline (n/H1100510) or various doses of R116301 (n/H110055/dose
`group). The individual scores are represented by small, white, vertical
`bars (5/dose group) on a black background. In this way, the degree of
`inhibition can be readily estimated from the visible area of black back-
`ground.
`700 Megens et al.
`Page 5 of 14
`
`
`
`
`
`
`
`Inhibition of SP-Induced Plasma Extravasation in Guinea
`Pigs
`Figure 2 shows individual scores (ranging from 0 to 3) for
`the SP-induced plasma extravasation in the nose, forepaws,
`and conjunctiva of guinea pigs 1 h after s.c. pretreatment
`with saline (n /H1100510) or various doses of R116301 (n /H110055/dose
`group). R116301 reduced the scores dose dependently for
`blue coloring in the three organs. The ED
`50 values for reduc-
`ing the scores below 2 were 0.097, 0.11, and 0.085 mg/kg for
`the nose, forepaws, and conjunctiva, respectively (Table 1).
`Table 1 compares R116301 with other NK
`1 receptor antag-
`onists at the indicated time interval after s.c., p.o., or i.v.
`administration. Regarding inhibition of extravasation in the
`nose after s.c. injection, R116301 (0.097 mg/kg) was 31 times
`less potent than GR-203040 (0.0031 mg/kg) but about equi-
`potent with aprepitant (0.028 mg/kg), nolpitantium (0.050
`mg/kg), CP-99994 (0.17 mg/kg), and CP-96345 (0.22 mg/kg).
`The p.o. over s.c. ED
`50 ratio was much better for R116301
`(2.7) and aprepitant (3.0) than for CP-96345 (12), CP-99994
`(124), GR-203040 (190), and the quaternary compound nol-
`pitantium (/H1102250).
`At 4 times the peak-effect dose (graphical estimated: 0.27
`mg/kg), R116301 showed a rapid onset (/H110211 h) and a relatively
`long duration of action (16 h; Fig. 3). In contrast, activity had
`already appreciably declined 4 h after s.c. injection of nolpi-
`tantium or p.o. administration of CP-96345 (Table 1). The
`rat-selective NK
`1 receptor antagonist RP-67580 hardly in -
`hibited extravasation in guinea pigs at the dose of 10 mg/kg,
`s.c. After an s.c. dose of 40 mg/kg, the NK
`2 receptor antago-
`nist saredutant (40 mg/kg, s.c.), the NK3 receptor antagonist
`osanetant (/H1102240 mg/kg, s.c.), and the histamine H1 receptor
`antagonists oxatomide and levocabastine did not at all affect
`the SP-induced plasma extravasation at doses far above the
`dose required for their primary activity.
`Skin Reaction Test in Guinea Pigs: SP, Histamine,
`PAF, and Bradykinin. R116301 (s.c., /H110022 h) was compared
`with the histamine H
`1 receptor antagonist levocabastine
`Fig. 3. ED50 (milligrams per kilogram) of R116301 for inhibition of
`SP-induced extravasation in the nose plotted versus time interval after
`oral administration. Vertical bars represent the 95% confidence limits to
`the ED
`50 values.
`TABLE 1
`ED50 values (95% CL; milligrams per kilogram) of R116301 and reference NK 1 receptor antagonists for inhibition of SP-induced extravasation in
`guinea-pigs
`Compound Route Time
`ED50 (95% C
`bis(Trifluoromethyl)benzoyl]-2-(phenylmethyl)-4-piperidinyl]-N-
`(2,6-dimethylphenyl)-1-acetamide (S)-Hydroxybutanedioate
`(R116301), an Orally and Centrally Active Neurokinin-1
`Receptor Antagonist
`A. A. H. P. MEGENS, D. ASHTON, J. C. A. VERMEIRE, P. C. M. VERMOTE, K. A. HENS, L. C. HILLEN,
`J. F. FRANSEN, M. MAHIEU, L. HEYLEN, J. E. LEYSEN, M. R. JURZAK, and F. JANSSENS
`Department of Discovery Research Johnson & Johnson Pharmaceutical Research and Development, Beerse, Belgium
`Received February 6, 2002; accepted April 9, 2002
`ABSTRACT
`In comparison with a series of reference compounds, (2R-trans)-
`4-[1-[3,5-bis(trifluoromethyl)benzoyl]-2-(phenylmethyl)-4-piperidi-
`nyl]-N-(2,6-dimethylphenyl)-1-acetamide (S)-Hydroxybutanedio-
`ate (R116301) was characterized as a specific, orally, and centrally
`active neurokinin-1 (NK
`1) receptor antagonist with subnanomolar
`affinity for the human NK1 receptor (Ki: 0.45 nM) and over 200-fold
`selectivity toward NK2 and NK3 receptors. R116301 inhibited sub-
`stance P (SP)-induced peripheral effects (skin reactions and
`plasma extravasation in guinea pigs) and a central effect (thump-
`ing in gerbils) at low doses (0.08–0.16 mg/kg, s.c. or i.p.), reflect-
`ing its high potency as an NK
`1 receptor antagonist and excellent
`brain disposition. Higher doses blocked various emetic stimuli in
`ferrets, cats, and dogs (ED
`50 values: 3.2 mg/kg, s.c.; 0.72–2.5
`mg/kg, p.o.). Even higher doses (11–25 mg/kg, s.c.) were required
`in mice (capsaicin-induced ear edema) and rats (SP-induced ex-
`travasation and salivation), consistent with lower affinity for the
`rodent NK
`1 receptor and known species differences in NK1 re-
`ceptor interactions. R116301 inhibited the ocular discharge (0.034
`mg/kg) but not the dyspnoea, lethality, or cough (/H1102240 mg/kg, s.c.)
`induced by [
`/H9252ALA8]-neurokinin A (NKA) (4–10) in guinea pigs,
`attesting to NK 1 over NK2 selectivity. R116301 did not affect
`senktide-induced miosis (/H110225 mg/kg, s.c.) in rabbits, confirming
`the absence of an interaction with the NK3 receptor. R116301 was
`inactive in guinea pigs against skin reactions induced by hista-
`mine, platelet-aggregating factor, bradykinin, or Ascaris allergens
`(/H1102210 mg/kg, s.c.). In all species, R116301 showed excellent oral
`over parenteral activity (ratio, 0.22–2.7) and a relatively long dura-
`tion (6.5–16 h, p.o.). The data attest to the specificity and sensi-
`tivity of the animal models and support a role of NK
`1 receptors in
`various diseases.
`Tachykinins belong to a family of short peptides that are
`widely distributed in the mammalian central and peripheral
`nervous system (Lundberg, 1995; Maggi, 1995; Bertrand and
`Geppetti, 1996). They share the common C-terminal se-
`quence Phe-Xaa-Gly-Leu-Met-NH
`2. Tachykinins released
`from peripheral sensory nerve endings are believed to be
`involved in neurogenic inflammation. In the spinal cord/cen-
`tral nervous system, tachykinins may play a role in pain
`transmission/perception and in some autonomic reflexes and
`behaviors. The three major tachykinins are substance P (SP),
`neurokinin (NK) A and NKB with preferential affinity for
`three distinct receptor subtypes, termed NK
`1,N K2, and NK3,
`respectively. However, functional studies on cloned receptors
`suggest strong functional cross-interaction between the three
`Article, publication date, and citation information can be found at
`http://jpet.aspetjournals.org.
`DOI: 10.1124/jpet.102.034348.
`ABBREVIATIONS:SP, substance P; NK, neurokinin; NKA, neurokinin A; R116301, (2R-trans)-4-[1-[3,5-bis(trifluoromethyl)benzoyl]-2-(phenylmethyl)-4-
`piperidinyl]-N-(2,6-dimethylphenyl)-1-acetamide ( S)-hydroxybutanedioate; CGP49823, (2 R,4S)-2-benzyl-1-(3,5-dimethylbenzoyl)-4-(quinolin-4-
`ylmethylamino)piperidine; CP-96345, (2S,3S)-cis-2-(diphenylmethyl)-N-((2-methoxyphenyl)-methyl)-1-azabicyclo(2.2.2.)-octan-3-amine; CP-99994, (2S,3S)-3-
`(2-methoxybenzylamino)-2-phenylpiperidine; GR-203040, (2S,3S)-(2-methoxy-5-tetrazol-1-ylbenzyl)(2-phenylpiperidin-3-yl)amine; MK-869 or L-754030,
`aprepitant; L-760,735, 2-(R)-(1-(R)-3,5-bis(trifluoromethyl)phenyl)ethoxy)-4-(5-(dimethylaminomethyl)-1,2,3-trioazol-4-yl)methyl-3-(5)-phenyl)morpholine; RP-
`67580, (3aR,7aR)-7,7-diphenyl-2-[1-imino-2-(2-methoxyphenyl)ethyl]perhydroisoindol-4-one; SDZ-NKT-343, 2-nitrophenylcarbamoyl-(S)-prolyl-(S)-3-(2-
`naphthyl)alanyl-N-benzyl-N-methylamide; SR-140333, nolpitantium; SR-48968, saredutant; SR-142801, osanetant; Y-24180, (/H11006)-4-(2-chlorophenyl)-2-[2-(4-
`isobutylphenyl)ethyl]-6,9-dimethyl-6H-thieno[3,2-f][1,2,4]triazolo[4,3-a][1,4]diazepine; LY255582, (3R,4R)-3,4-dimethyl-1-[(3S)-3-hydroxy-3-cyclohexyl-propyl]-
`4-(3-hydroxyphenyl)piperidine; PAF, platelet-aggregating factor; 5-HT, serotonin; MDL-103392, 4-piperidinecarboxamide, 1-[2-[3-(3,4-dichlorophenyl)-1-(3,4,5-
`trimethoxybenzoyl)-3-pryrrolidinyl]ethyl]-4-phenyl; MDL-105212, (3R)-MDL-103392.
`0022-3565/02/3022-696–709$7.00
`THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 302, No. 2
`Copyright © 2002 by The American Society for Pharmacology and Experimental Therapeutics 34348/996964
`JPET 302:696–709, 2002 Printed in U.S.A.
`696
`HELSINN EXHIBIT 2066
`Azurity Pharmaceuticals, Inc. v. Helsinn Healthcare S.A.
`IPR2025-00948
`Page 1 of 14
`
`
`
`
`
`
`
`tachykinins and their corresponding receptors (Maggi and
`Schwartz, 1997). The tachykinins are involved in emesis,
`anxiety states (stress-related), smooth muscle contraction,
`inflammation, and nociception/pain perception, although
`clinically relevant involvement of SP, particularly for the last
`items, is questionable (Hill, 2000).
`R116301 is a new NK
`1 receptor antagonist (Fig. 1 ). The
`present study reports on its pharmacological profile in sev-
`eral NK receptor-related models in various species. Species
`differences in the structure of NK
`1 receptors are responsible
`for species-related potency differences of NK1 receptor antag-
`onists (Maggi, 1995). The human NK1 receptor closely resem-
`bles the NK 1 receptor of guinea pigs and gerbils but differs
`markedly from the NK1 receptor of rodents. Therefore, most
`tests are performed in guinea pigs and gerbils. Attention is
`paid to potency, selectivity, specificity, onset and duration of
`action, and species differences. R116301 is compared with
`the following available NK
`1 receptor antagonists (Fig. 1 for
`chemical structures): CGP49823 (Vassout et al., 1994), CP-
`96345 (Snider et al., 1991), CP-99994 (Piedimonte et al.,
`1993), GR-203040 (Ward et al., 1995), MDL-103392 (race-
`mate of the active enantiomer MDL-105212; Kudlacz et al.,
`1996), aprepitant (MK-869 or L-754030; Kramer et al., 1998;
`Rupniak and Kramer, 1999), L-760735 (McAllister et al.,
`Fig. 1. Chemical structure of R116301 [(S)-hydroxybutanedioate salt] and the reference NK 1 receptor antagonists.
`Pharmacology of R116301 697
`Page 2 of 14
`
`
`
`
`
`
`
`1999), RP-67580 (Garret et al., 1991), SDZ-NKT-343 (Wal-
`pole et al., 1998), nolpitantium (SR-140333; Emonds-Alt et
`al., 1993). The testing of some compounds was restricted by
`limited availability. The sensitivity, reliability, and specific-
`ity of the various test models is evaluated based on the
`results obtained with the test compounds. A preliminary
`article on R116301 has been presented in poster format
`(Jurzak et al., 2000). A patent application has been filed
`(Janssens et al., 1997).
`Materials and Methods
`Animals
`Swiss mice (Janssen Pharmaceutica, Beerse, Belgium), Wistar
`rats (Janssen Pharmaceutica), Mongolian CRW gerbils ( Meriones
`unguiculatus; Charles River Breeding Laboratories, Inc., Sulzteld,
`Germany), Dunkin-Hartley-Purbright guinea pigs (Janssen Phar-
`maceutica or Charles River Breeding Laboratories, Inc.), Fish ferrets
`(Harlan CPB, Horst, The Netherlands), New Zealand white rabbits
`(Broekman Institute, Someren, The Netherlands), cats (Broekman
`Institute), and Beagle dogs (Janssen Pharmaceutica or Harlan CPB)
`were used. They were fasted overnight (tap water remained avail-
`able ad libitum) and housed under standard laboratory conditions
`(21 /H110062°C; 65 /H1100615% relative humidity; light/dark cycle set at 12 h).
`During the test period, they were housed in individual cages. The
`ferrets, cats, and dogs were used more than once, with an intertrial
`interval of at least 1 week. All animal studies were approved by the
`local Ethical Committee in compliance with the Declaration of Hel-
`sinki.
`Chemicals and Test Compounds
`R116301 was dissolved up to 2.5 mg/ml in 10% hydroxypropyl- /H9252-
`cyclodextrin. Higher doses were prepared as suspensions in 1% poly-
`sorbate 80 in distilled water. The preparations were stored at room
`temperature in closed containers protected from light. They were
`studied at various time intervals after subcutaneous (10 ml/kg for
`mice, rats, and guinea pigs; 1.0 ml/kg for rabbits; 0.5 ml/kg for cats
`and dogs), oral (10 ml/kg for mice, rats, and guinea pigs; 1.0 ml/kg for
`rabbits; 0.5 ml/kg for cats and dogs), or intravenous administration
`(10 ml/kg for mice; 2 ml/kg for rats; 0.5 ml/kg for rabbits, cats, and
`dogs). All doses were expressed in milligram base equivalents per
`kilogram body weight. With the exception of aprepitant (L-754030 or
`MK-869; synthesized by and obtained from Johnson & Johnson
`Pharmaceutical Research Institute and Development, Springhouse,
`NJ) and L-760735 (synthesized by our Department of Medicinal
`Chemistry), the reference compounds were kindly provided by the
`companies of origin: CGP49823 and SDZ-NKT-343 (Novartis, Basel,
`Switzerland); CP-96345 and CP-99994 (Pfizer, Sandwich, Kent, UK);
`GR-203040
`(GlaxoSmithKline, Uxbridge, Middlesex, UK); MDL-103392 (Aven-
`tis, Strasbourg, France); RP-67580 (Aventis); saredutant/SR-48968,
`nolpitantium/SR-140333, and osanetant/SR-142801 (SANOFI Re-
`search Center, Montpellier, France). For all these compounds with
`exception of SR-140333 and SDZ-NKT-343, some batches were syn-
`thesized in our Department of Medicinal Chemistry. The origin of
`the other chemicals is indicated in parentheses after each name:
`substance P (Sigma-Aldrich, St. Louis, MO), [
`/H9252ALA8]-neurokinin A
`(4–10) (NovaBiochem, Laufelfingen, Switzerland), capsaicin (Sigma-
`Aldrich), PAF (Sigma-Aldrich), bradykinin (Sigma-Aldrich), hista-
`mine (Sigma-Aldrich), and Evans blue dye (Sigma-Aldrich; Direct
`blue 53; dye content approximately 85%). Ascaris suum worms were
`obtained from freshly slaughtered pigs. A batch of about 20 ml of
`perienteric fluid was centrifuged; the clear supernatant was divided
`in 0.1-ml portions and stored at /H1100218°C. The injected solutions of
`Ascaris coeloma fluid were fresh 1:16 dilutions in 0.9% NaCl.
`Pharmacological Tests
`In Vitro Receptor Binding. Membrane preparation from cells.
`Chinese hamster ovary cells expressing human (h)NK1,N K2 or NK3
`receptors were grown in Dulbecco’s modified Eagle’s medium/HAM’s
`F-12 medium containing 10% fetal calf serum and antibiotics. Twen-
`ty-four hours after stimulation with 5 mM Na-butyrate to enhance
`expression levels, the cells were collected from plates using a rubber
`scraper and suspended in 50 mM Tris-HCl, pH 7.4. They were
`centrifuged at 23,500g for 10 min in a Sorvall-RC 5B centrifuge
`(DuPont Instruments, Meyvis, Belgium). The pellets were homoge-
`nized in 5 mM Tris-HCl, pH 7.4, using an Ultra-Turrax homogenizer
`(Janke & Kunkel IKA Labortechnik, Staufen im Breisgau, Germany)
`and centrifuged at 30,000g for 20 min. The final pellet was sus-
`pended in 50 mM Tris-HCl, pH 7.4. The membranes were frozen in
`1-ml aliquots /H1100270°C. Before use, vials were thawed and rehomog-
`enized in incubation buffer B (50 mM Tris-HCl, pH 7.4, containing 2
`mM MgCl
`2, 1 mM EGTA, and 0.1% bovine serum albumin). The
`protein concentration was determined after using a Bradford kit
`from Bio-Rad (Hercules, CA).
`Membrane preparation from brain tissue. To prepare membranes
`for [
`3H]substance P binding from guinea pigs, gerbils, ferrets, and
`rats, animals were killed by decapitation, and the forebrains were
`dissected. The tissue was homogenized in 50 mM Tris-HCl, pH 7.4,
`using an Ultra-Turrax homogenizer. The homogenates were centri-
`fuged at 23,500g for 10 min at 0 –4°C in a Sorvall-RC 5B centrifuge.
`The pellets were washed twice in 50 mM Tris-HCl, pH 7.4, by
`resuspension with a dual-homogenizer (Kimble Kontes, Vineland,
`NJ) and centrifugation. After the last wash, the pellets were sus-
`pended in incubation buffer A [50 mM Tris-HCl, pH 7.4, containing
`2 mM MgCl
`2, 1 mM EGTA, 1 mM o-phenantrolin, and 0.1% bovine
`serum albumin at a dilution of 25 volumes/original wet weight of
`tissue (v/w)]. The final pellet of rat forebrain was suspended in
`incubation buffer B at a dilution of 40 v/w. Guinea pig, gerbil, and rat
`forebrain membranes were always freshly prepared. For ferret fore-
`brain, the membranes were prepared in 50 mM Tris-HCl, pH 7.4,
`and stored at a dilution of 5 v/w at /H1100270°C. Before use, these mem-
`branes were thawed and further diluted until dilution of 25 v/w in
`incubation buffer A.
`[
`3H]Substance P binding to NK1 receptors and data analysis. The
`membrane homogenates were incubated for 20 min at 25°C with 0.5
`nM [
`3H]SP (for rat membranes 1 nM was used) in a total volume of
`0.5 ml. Specific binding of the radioligand was distinguished from
`nonspecific binding by addition of unlabeled SP to a final concentra-
`tion of 10
`/H110027M. The incubation was stopped by the addition of 5 ml of
`ice-cold 50 mM Tris-HCl buffer, pH 7.4, followed by rapid filtration
`over Whatman GF/B glass fiber filters (Maidstone, Kent, UK) (pre-
`soaked in 0.1% polyethylenimine for 1 h) using a 40-well filtration
`unit. The filters were washed twice with ice-cold buffer to remove
`nonbound radioactivity and placed in plastic miniature vials. After a
`24-h incubation with 2 ml of Ultima Gold scintillation cocktail, vials
`were vigorously shaken, and the radioactivity was counted in a
`Packard Tri-Carb 1500 CA liquid scintillation analyser (Packard
`BioScience, Meriden, CT).
`R116301 was also investigated in various other in vitro binding
`assays using membrane preparations of animal tissue or membranes
`of cell lines transfected with cloned human receptors. Brain, periph-
`eral organs, blood of animal or human origin, or permanent cell lines
`were used as tissue sources. The procedures for membrane prepara-
`tions and references to the various receptor binding models were
`described previously (Briejer et al., 2001).
`Substance P-Induced Plasma Protein Extravasation in
`Guinea Pigs. SP (2
`/H9262g/kg, i.v.) and Evans blue dye (30 mg/kg, i.v.)
`were injected simultaneously (one solution; 4 ml/kg) into the femoral
`artery of guinea pigs of both sexes (325– 425 g) at predefined, loga-
`rithmically spaced, time intervals (1, 2, 4, 8, 16 or 32 h) after
`pretreatment with test compound or solvent. Up to 10 min after
`challenge, the animals were scored by visual inspection for blue
`698 Megens et al.
`Page 3 of 14
`
`
`
`
`
`
`
`coloring (0, 1, 2, or 3) of the nose, the forepaws, and the conjunctiva.
`The criterion for drug-induced inhibition of plasma extravasation:
`score /H110212 (occurrence in 0.2, 0.0, and 0.0% for blue coloring of the
`nose, paws, and conjunctiva, respectively, in saline-treated controls;
`n /H11005500).
`Capsaicin-Induced Ear Edema in Mice. Ear edema was in-
`duced by local application of capsaicin (250
`/H9262g) on the ear of female
`mice (20 –26 g) pretreated with test compound or solvent (modified
`after Inoue et al., 1996). The capsaicin was dissolved in acetone at a
`concentration of 12.5 mg/ml and applied in a volume of 10
`/H9262l both on
`the ventral and dorsal side of the left ear. Pure acetone was applied
`in a volume of 10
`/H9262l both on the ventral and dorsal side of the right
`ear, which served as a control measure. Immediately after the cap-
`saicin challenge, Evans blue dye (7.5 mg/ml in 0.9% NaCl) was
`injected intravenously in a volume of 10 ml/kg, corresponding to a
`dose of 75 mg/kg. Thirty minutes after the capsaicin application, ear
`edema was scored (0, 1, 2, or 3) as the blue coloring of the left ear.
`Criterion for drug-induced inhibition: score /H110212 (never observed in
`controls; n /H1100562).
`Substance P-Induced Salivation and Plasma Protein Ex-
`travasation in Rats. Plasma protein extravasation and salivation
`were induced by injection of SP (2
`/H9262g/4 ml/kg, i.v.) in the tail vein of
`female rats (200 –240 g) that were anesthetized with pentobarbital
`(40 mg/kg, i.p.; 10 min before the SP challenge) and placed on a
`heating pad to maintain body temperature [modified after Snider et
`al. (1991); Robineau et al. (1995)]. Evans blue dye (30 mg/kg, i.v.) was
`injected simultaneously with SP (one solution; 4 ml/kg). Test com-
`pound or solvent was administered at a predefined time interval
`before SP injection. From 0 through 4 min after the SP challenge,
`salivation was measured by placing cotton swabs in the rat ’s mouth
`at 2-min intervals and quantifying the amount of saliva secreted by
`the difference in the weight of the two cotton swabs before and after
`the collection period. Five minutes after challenge, the animals were
`scored for blue coloring (0, 1, 2, or 3) of the nose and the paws. Thirty
`minutes after challenge, the animals were sacrificed by CO
`2 asphyx-
`iation and scored (0, 1, 2, or 3) for blue coloring of the trachea and
`urinary bladder. Criteria for drug-induced inhibition: /H11021100 mg of
`saliva for inhibition of salivation (2.0% in controls;n /H11022100); score /H110212
`for inhibition of plasma extravasation (0, 0, 1.0, and 1.0 for blue
`coloring of the nose, paws, trachea, and urinary bladder, respec-
`tively, in controls).
`Skin Reaction Test in Guinea Pigs: SP, Histamine, PAF,
`Bradykinin, and Ascaris Allergens. Five dorsal skin sites of
`guinea pigs of both sexes (300 –500 g) were injected intradermally
`(0.05 ml) with saline containing SP (0.05
`/H9262g), histamine (0.25 /H9262g),
`PAF (0.00625 /H9262g), bradykinin (0.1 /H9262g), and Ascaris allergens (1/16
`diluted with saline). Immediately thereafter, the animals were chal-
`lenged i.v. with Evans blue dye (30 mg/kg; 7.5 mg/ml, 4 ml/kg) and
`sacrificed 30 min later. The intensity of the blue-colored skin reac-
`tions was scored by two independent observers in comparison with
`standard reactions. The scoring system was maximal (4), pronounced
`(3), moderate (2), slight (1), and no difference (0) with surrounding
`skin. The scores of the two observers were summed for further
`evaluation. Test compound or solvent was administered at a pre-
`defined interval before induction of the skin reactions. Criteria for
`drug-induced effects were: 1) SP reactions: score /H110217 for inhibition,
`score /H110215 for pronounced inhibition, and score/H110213 for blockade (oc-
`curring in 3.6, 0.0, and 0.0%, respectively, of solvent-treated control
`guinea pigs; n /H1100557); 2) histamine reactions: score /H110217 for inhibition
`and score /H110213 for blockade (occurring in 3.6% and 0.0%, respectively,
`of the controls); 3) PAF reactions: score /H110217 for inhibition and score
`/H110213 for blockade (both occurring in 0.0% of the controls); and 4)
`bradykinin reactions: score /H110217 for inhibition and score/H110213 for block-
`ade (both occurring in 0.0% of the controls); and 5) Ascaris allergens
`reactions: score /H110213 for blockade (occurring in 5.4% of the controls).
`SP-Induced Thumping in Gerbils. Male gerbils (50–70 g) were
`prepared for intracerebral injection under anesthesia (exposure to
`4% isoflurane in 70% N
`2O /H1100130% O2 during 1 min) by making an
`incision in the skin above the cranium (modified after Bristow and
`Young, 1994 and Rupniak et al., 1997). SP (400 ng in 2 /H9262l) was
`rapidly injected into the cerebral ventricles after advancing a Ham-
`ilton needle 3.5 mm below a point 2 mm anterior to bregma and 1 mm
`lateral to the midline; the needle was removed 10 s later. The
`animals were observed for thumping behavior during the following 5
`min. Thumping was defined as a sharp downward movement of the
`hind-paws producing a characteristic rhythmic tapping sound. The
`onset of the SP-induced thumping, the total number of thumps, and
`the total thumping time (seconds) were registered. The animals were
`then immediately euthanized with CO
`2. For the present purpose,
`absence of thumps during the 5-min observation period was adopted
`as an all-or-nothing criterion for inhibition of the SP-induced thump-
`ing (1.2% false positive controls; n /H11005625).
`[
`/H9252ALA8]-NKA (4–10)-Induced Lethality in Guinea Pigs. The
`following phenomena were scored or noted after injection of [/H9252ALA8]-
`neurokinin A (4 –10) (50 /H9262g/4 ml/kg; i.v.) into the vena saphena of
`female guinea pigs (300 –500 g) pretreated with test compound or
`solvent: cough (intensity scores: 0, 1, or 2), white ocular discharge
`(scores: 0, 1, or 2), dyspnoea (score 1 /H11005cyanosis, score 2 /H11005dyspnoea,
`score 3 /H11005loss of righting reflex and/or clonic convulsions), and
`lethality (the survival time up to 60 min after [
`/H9252ALA8]-NKA (4 –10)
`challenge is noted) (modified after Robineau et al., 1995). [ /H9252ALA8]-
`neurokinin A (4 –10) is a metabolism-resistant analog of neuroki-
`nin-A with a high degree of selectivity for the NK 2 receptor (Rovero
`et al., 1989). The various phenomena generally occurred within 3 to
`5 min after the [
`/H9252ALA8]-NKA (4–10) challenge. The following all-or-
`nothing criteria were adopted for the determination of the ED 50
`values of the test compounds: score /H110050 for antagonism of cough
`(occurrence: 0.0% false positives in the control population; n /H1100596),
`score /H110050 for antagonism of ocular discharge (2.1% false positives),
`score /H110212 for protection from dyspnoea (1.0% false positives), and
`survival time /H1102260 min for protection from lethality (5.2% false pos-
`itives).
`Emesis in Ferrets, Cats, and Dogs. Vomiting or retching was
`induced by the peripherally selective opioid loperamide (0.31 mg/kg,
`s.c.), by ipecac syrup (1 ml/kg, p.o.; from a commercial source and
`containing ether-soluble alkaloids of ipecac in a concentration of 1.4
`mg/ml), by the dopamine agonist apomorphine (0.31 mg/kg, s.c.), or
`by the
`/H92512-adrenoceptor agonist xylazine (1.25 mg/kg, s.c.) in male
`ferrets (1–3 kg), cats (3–7 kg), or dogs (5–20 kg) pretreated with test
`compound or solvent. The number of retches or vomits was counted
`over a 1 h-period or (in the case of ipecac syrup-induced emesis in
`cats) a 3 h-period starting immediately after the emetic challenge.
`Loperamide-induced retching in ferrets. In control animals pre-
`treated with solvent, loperamide (0.31 mg/kg, s.c.) induced pro-
`nounced retching (mean /H11006S.D.: 95 /H1100639 counts; n /H11005529) and, to a
`lesser extent, vomiting (5 /H110064). All-or-nothing criteria for drug-
`induced effects on retching: /H1102120 retches for inhibition (2.0% false
`positives) and 0 retches for blockade (0% false positives).
`Ipecac syrup-induced retching in ferrets. Oral administration of
`ipecac syrup induced pronounced retching (mean /H11006S.D.: 76 /H1100636
`counts) and, to a lesser extent, vomiting (mean /H11006S.E.: 5.6 /H110062.3
`counts; n /H1100598) in control animals. Absence of retching or vomiting
`never occurred in these control animals and was used as criterion for
`antiemetic activity.
`Apomorphine-induced retching in ferrets.Injection of apomorphine
`resulted in pronounced retching in control ferrets (mean/H11006S.D.: 63 /H11006
`29 counts; n /H1100592) and, to a lesser extent, vomiting (mean /H11006S.D.:
`3.1 /H110062.0 counts). Absence of retching never occurred in these control
`animals and was used as criterion for antiemetic activity. Vomiting
`was absent in 13% of the control animals.
`Apomorphine-induced vomiting in dogs. Apomorphine induced
`consistent vomiting in all control dogs (mean /H11006S.D.: 10 /H110065 vomits;
`n /H11005195). Criteria for drug-induced protection: inhibition of emesis:
`/H110212 emetic bouts (occurring in 0.12% of controls); blockade of emesis:
`complete absence of emesis (not observed in controls).
`Xylazine-induced retching in cats. Control cats (n /H11005113) retched
`Pharmacology of R116301
`699
`Page 4 of 14
`
`
`
`
`
`
`
`(mean /H11006S.D.: 41 /H1100618 counts) and vomited (mean /H11006S.D.: 2.9 /H110061.8
`counts) after injection of xylazine. Absence of retching or vomiting
`never occurred and was used as criterion for antiemetic activity.
`Ipecac syrup-induced vomiting in cats. Oral administration of ip-
`ecac syrup induced vomiting and retching during a 3-h observation
`period in control cats pretreated with distilled water (n /H1100530). For
`practical reasons, only the number of emetic bouts was counted,
`which remained relatively low (mean /H11006S.E.: 4.5 /H110060.6 counts).
`Absence of vomiting never occurred and was used as the criterion for
`antiemetic activity.
`Senktide-Induced Miosis in Rabbits. Senktide (12.5
`/H9262g/0.5
`ml/kg, i.v.)-induced miosis was evaluated just before and 15 min
`after challenge in male New Zealand white rabbits (approximately 1
`kg) pretreated with test compound or solvent (1.0 ml/kg) (modified
`after Medhurst et al., 1997). The senktide was injected into an ear
`vein. The left pupil of each rabbit was measured under normal
`ambient lighting with a comparing reticule (scaled to 0.5 mm). In
`solvent-pretreated control rabbits (n /H1100573), pupil diameter declined
`from 6.0 /H110060.5 mm immediately before to 2.7 /H110060.7 mm 15 min after
`the senktide challenge. At this 15-min time interval after senktide
`injection, only two of the control rabbits (2.7%) showed a pupil
`diameter /H113505.0 mm, which was adopted as all-or-nothing criterion for
`significant inhibition of the senktide-induced miosis by test com-
`pounds.
`General Procedure and Statistics
`In Vitro Binding Studies. Sigmoidal inhibition curves were
`calculated by computerized nonlinear regression analyses according
`to Oestreicher and Pinto. pIC
`50 values (/H11002log concentration producing
`50% inhibition of specific [ 3H]SP binding) were derived. Inhibition
`equilibrium constants (K i values) were calculated according to Ki
`(nanomolar) /H11005IC50/(1 /H11001C/Kd), where C is the concentration and Kd
`the equilibrium dissociation constant of the [ 3H]SP.
`In Vivo Functional Studies.All experiments were performed by
`unbiased trained technicians using coded solutions. Doses were se-
`lected from the geometrical series 0.00063–0.00125–0.0025. . . 40.0–
`80.0–160 mg/kg in such a way that at least three doses covered the
`0 to 100% effect range of the dose-response curve. Each dose group
`consisted of five animals, which were tested in separate daily exper-
`imental sessions (including solvent-treated control animals) to ac-
`count for day-to-day variability and to minimize systematic errors.
`Control injections of solvent were included in each experimental
`session. Based on an analysis of a frequency distribution of a large
`series of the historical control data, all-or-nothing criteria for stim-
`ulation or inhibition were defined as a change of the measured or
`scored variable to values that never or almost never (in less than 5%)
`occurred in the control population. With the help of the thus defined
`all-or-nothing criteria, graded data were transformed to categorical
`data which generated per dose level the number of animals in which
`the intensity of a particular phenomenon was higher (stimulation) or
`lower (inhibition or blockade) than in the control animals. On the
`basis of the thus obtained dose-response relations, ED
`50 values and
`corresponding 95% confidence limits were calculated by probit anal-
`ysis according to the method of Finney (1962) for categorical data.
`Results
`In Vitro Receptor Binding Profile
`R116301 showed subnanomolar affinity for the human
`NK1 receptor (pIC50 /H11006S.D.: 8.53 /H110060.06; Ki: 0.45 nM; n /H110054
`replicates), with 230- and 1600-fold selectivity regarding
`binding to the human NK3 (pIC50 /H11006S.D.: 6.22 /H110060.14; Ki: 104
`nM; n /H110053) and NK2 (pIC50 /H11006S.D.: 5.87 /H110060.36; Ki: 711 nM;
`n /H110054) receptors, respectively. R116301 showed very low
`affinity for human serotonin 5-HT 2B (pIC50 /H11006S.D.: 5.68 /H11006
`0.15; Ki: 928 nM;n /H110052) and 5-ht5a (pIC50 /H11006S.D.: 5.16 /H110060.04;
`Ki: 3544 nM; n /H110052) receptors and for rat Ca2/H11001ligand binding
`sites (pIC50 /H11006S.D.: 5.45 /H110060.15; Ki: 2454 nM; n /H110053) and Na/H11001
`channels (pIC50 /H11006S.D.: 5.60 /H110060.17; Ki: 2526 nM; n /H110053). Up
`to 10 /H9262M, the compound had no affinity for a wide range of
`other binding sites, including adrenergic receptors (human
`/H92511A-, /H92512A-, /H92512B-, /H92512C-, /H92521-, /H92522-, /H92523-), dopaminergic (rat D 1;
`human D 2L,D 3, and D 4), serotonergic (human 5-HT 1A,
`5-HT1B, 5-HT 1D, 5-ht 1e, 5-ht 1f, 5-HT 2A, 5-HT 2C, 5-HT 3,
`5-HT4b, and 5-HT 7), human histamine H 1, rat cholinergic
`muscarinic, transporters (rat DA, NE; human 5-HT, GlyT 1,
`and GlyT 2), aspartate (rat NMDA-MK801, NMDA-glycine,
`and AMPA), opioid (human /H9262, /H9254; guinea pig /H9260), human halo-
`peridol-sensitive /H92681, peptide receptors (rat CCK A; human
`CCKB, bradykinin-B2, VIP). Since tachykinin receptors are
`known to exhibit strong species dependence in pharmacology,
`the NK1 affinity of R116301 was investigated on tissue mem-
`brane preparations from forebrains of guinea pigs, gerbils,
`ferrets, and rats. Relative to its affinity for the human NK
`1
`receptor, R116301 showed about 10 times lower affinity for
`the gerbil, ferret, and guinea pig NK
`1 receptors (Ki: 6.4, 8.3,
`and 13 nM, respectively) and 200 times lower affinity for the
`rat NK
`1 receptor (K i: 98 nM). These results show a high
`affinity, selectivity, and specificity of R116301 for the human
`NK
`1 receptor and a species preference as generally observed
`for other NK1 receptor antagonists as well.
`Fig. 2. Individual scores (ranging from 0 to 3) for SP-induced blue
`coloring of the nose, forepaws, and conjunctiva of guinea pigs 1 h after s.c.
`pretreatment with saline (n/H1100510) or various doses of R116301 (n/H110055/dose
`group). The individual scores are represented by small, white, vertical
`bars (5/dose group) on a black background. In this way, the degree of
`inhibition can be readily estimated from the visible area of black back-
`ground.
`700 Megens et al.
`Page 5 of 14
`
`
`
`
`
`
`
`Inhibition of SP-Induced Plasma Extravasation in Guinea
`Pigs
`Figure 2 shows individual scores (ranging from 0 to 3) for
`the SP-induced plasma extravasation in the nose, forepaws,
`and conjunctiva of guinea pigs 1 h after s.c. pretreatment
`with saline (n /H1100510) or various doses of R116301 (n /H110055/dose
`group). R116301 reduced the scores dose dependently for
`blue coloring in the three organs. The ED
`50 values for reduc-
`ing the scores below 2 were 0.097, 0.11, and 0.085 mg/kg for
`the nose, forepaws, and conjunctiva, respectively (Table 1).
`Table 1 compares R116301 with other NK
`1 receptor antag-
`onists at the indicated time interval after s.c., p.o., or i.v.
`administration. Regarding inhibition of extravasation in the
`nose after s.c. injection, R116301 (0.097 mg/kg) was 31 times
`less potent than GR-203040 (0.0031 mg/kg) but about equi-
`potent with aprepitant (0.028 mg/kg), nolpitantium (0.050
`mg/kg), CP-99994 (0.17 mg/kg), and CP-96345 (0.22 mg/kg).
`The p.o. over s.c. ED
`50 ratio was much better for R116301
`(2.7) and aprepitant (3.0) than for CP-96345 (12), CP-99994
`(124), GR-203040 (190), and the quaternary compound nol-
`pitantium (/H1102250).
`At 4 times the peak-effect dose (graphical estimated: 0.27
`mg/kg), R116301 showed a rapid onset (/H110211 h) and a relatively
`long duration of action (16 h; Fig. 3). In contrast, activity had
`already appreciably declined 4 h after s.c. injection of nolpi-
`tantium or p.o. administration of CP-96345 (Table 1). The
`rat-selective NK
`1 receptor antagonist RP-67580 hardly in -
`hibited extravasation in guinea pigs at the dose of 10 mg/kg,
`s.c. After an s.c. dose of 40 mg/kg, the NK
`2 receptor antago-
`nist saredutant (40 mg/kg, s.c.), the NK3 receptor antagonist
`osanetant (/H1102240 mg/kg, s.c.), and the histamine H1 receptor
`antagonists oxatomide and levocabastine did not at all affect
`the SP-induced plasma extravasation at doses far above the
`dose required for their primary activity.
`Skin Reaction Test in Guinea Pigs: SP, Histamine,
`PAF, and Bradykinin. R116301 (s.c., /H110022 h) was compared
`with the histamine H
`1 receptor antagonist levocabastine
`Fig. 3. ED50 (milligrams per kilogram) of R116301 for inhibition of
`SP-induced extravasation in the nose plotted versus time interval after
`oral administration. Vertical bars represent the 95% confidence limits to
`the ED
`50 values.
`TABLE 1
`ED50 values (95% CL; milligrams per kilogram) of R116301 and reference NK 1 receptor antagonists for inhibition of SP-induced extravasation in
`guinea-pigs
`Compound Route Time
`ED50 (95% C

